
| The migmatites of the
Kanmantoo Group
exposed in at Point Ellen at
the western end of Vivonne Bay, and along the coast towards the SW show
many features characteristic of melt extraction during folding (see
Foden et al., 2002, J.Geol. Soc. London, v. 159, 2002,
557-575).
My main findings were that wide axial planar leucosomes (dykes) with
numerous fragments of coutnry rock, trending 060-080 represent melt
escape pathways. These leucosomes played a key role in strain
accommodation by allowing development of disharmonic metric to
decametric folds. Their role during folding was partly to remove mass
from limbs, and partly to provide shear planes or planes of detachment,
allowing limbs to collapse or to rotate. Here is a list of some of my
findings. This list is supported by the figures below. 1. Leucosomes and dykes (referred to here generally as leucosomes) are parallel to the axial plane of metric and decametric open folds trending 060 to 090 and dipping 40 to 90 north (Fig. 1). Leucosomes are also found parallel to bedding, S0, defining the folds. 2. These ENE-trending folds have gently dipping limbs and generally define open folds, with a small interlimb angle (less than 90 degrees). They overprint an early fold phase, F1, evidenced by intrafolial, isoclinal folds (Fig. 8), and a strong north-trending lineation that is folded around the ENE trending open folds, F2. 2. Axial planar leucosomes include numerous fragments of partially molten, refractory and restitic material from its source forming a breccia (Fig. 2). Layer-parallel leucosomes do not include fragments, suggesting that the former are melt escape zones that mobilized the source, and the latter are more or less in situ. 3. Axial planar leucosomes are up to 2m wide and continuous for only a few metres to a maximum of ten metres and end by breaking up into a network of thin axial planar leucosomes (Fig. 4). 4. Axial planar leucosomes either truncate abruptly the gently dipping S0 or rotate it / transpose it (Fig. 1 and 3) defining normal shear zones along which bedding is rotated into parallelism with the axial planar foliation (Fig. 1). Commonly one side of the axial planar leucosome is truncated and the other is parallel to it. Here, I use to term normal shear zone to describe the fact that layers are extended across it, rather than repeated across it. AXIAL PLANAR LEUCOSOMES DISRUPTED ROCK CONTINUITY ALLOWING BLOCK ROTATION AND SHEARING 5. Parasitic Folds in the presence of melt have a characteristic step shape (Fig. 5), bu cuspate fold hinges (as described in Tangtse, Ladakh) are also found locally (Fig. 6). 6. The open F2 folds have a significant change in shape forming decametric-scale disharmonic folds, always associated with the presence of thick axial planar leucosomes. In these regions, the hinge zone is preserved as gently dipping areas, and the limbs are sharply and abruptly rotated along leucosome-filled axial planar shear zones. THIS IS THE MOST IMPORTANT FINDING because they define the collapse of folds caused by volume loss due to melt extraction. The thick leucosomes are part and parcel of fold collapse, where the open, undulating folds become disharmonic and the limbs rotate abruptly and steepen up due to mass loss along the leucosomes. It is an impossible geometry unless volume loss is considered. Axial planar melt remnants record regions of prefential melt extraction from the migmatites and volume loss which accommodated the disharmonic folds and truncation of fold limbs Structural question: I have linked the isoclinal folds with the strong intersection and stretching N-trending lineation and called them D1. Could there be more then one deformation phase overprinted by the ENE-trending opne F2 folds? Migmatites question: There were two separate migmatization events, and I have not sorted out their relationship. There is an early formed pegmatite that is cross-cut by later, ENE-trending axial planar pegmatitic to granitic leucosomes/dykes. This is most clear to the SW of Point Ellen. Early, dominantly layer parallel pegmatitic leucosomes are cut by later ENE-trending leucosomes (see Fig. 7). |
| Figure 1. These figures illustrate the most significant relationships between axial planar leucosomes and bedding. Commonly one side of the leucosome truncates S0 whereas the other rotates S0 into parallelism in a normal shear zone, which extends layers. |
 A: On the left, bedding (357/42W) is truncated by the axial planar leucosome (dyke) 065/80N. On the right hand side, bedding defines a drag fold rotating from parallel to the leucosome in its vicinity to 037/46 NW outwards. |
 B: Detail of A, from above. Note the fragments of country rock in the dyke. |
 C: On the left, bedding dipping W (into the plane of the photo) is gently dragged towards the axial planar leucosome (trending 070 and dipping moderately N). On the right hand side bedding is parallel to the leucosome. Drag folds define an extensional movement. |
 D: Axial planar leucosome (070-080/45N) truncating bedding on the left-hand side (S0=320/30SW), and truncating bedding with a different orientation on the right-hand side (S0=050/40NW). In this case there was no drag folding but an abrupt block rotation across the leucosome. |
 E: Strongly stretched ductile breccia (075/30NW) with narrow leucosomes defining blocks, truncating bedding (S0=350/75W) and an early formed pegmatite. Interpretation: this represents a melt-filled leucosome from which most of the melt has been extracted, leaving the country rock fragments. This transposition zone is 60 cm wide and breaks up into a number of narrow axial planar shear bands related to folds (F). The break-up of the transposition zone is gradual. |
 F: Along strike from E, bedding gently folded and cross-cut by narrow axial planar leucosomes separating folds. The change from E to F. |
| Figure 2. Breccias in axial planar leucosomes. |
 A: |
 B: |
 C: Breccia |
 D: Leucosome breccia trending 070 and cutting abruptly bedding (S0=345/30W). |
 E: Rotational flow around a pegmatite block. |
 F: Disruption of layered psammite. |
| Figure 3. Transposition of earlier layering by axial planar foliation with melt. |
 A: Transpoistion of layering due to melt migration. Further examples of transposition can be seen in many of the other photos on this page. |
| Figure 4 Merging of leucosome network. |
 A: Merging of diffuse, disorganized network of leucosomes into a broader dyke to the left |
 B: Same |
 C: Same as Fig. 6a, showing the merging of layer parallel leucosomes into an axial planar leucosome. |
| Figure 5. Typical step parasitic fold style in the presence of axial planar melt. |
 A: Step-folding characterisitic of folding in the presence of axial planar leucosomes linked with layer parallel leucosomes. |
 B: |
 C: |
| Figure 6. Cuspate folds indicating disruption caused by melt migration. Interpretation: melt moved upwards on photo, in a manner similar to cuspate folds. |
 A: Layer-parallel leucosomes rotating into and merging in the direction of the axial planar foliation (E-W on photo) forming a cuspate fold with a tip pointing to the left on photo (SW). Melt migration is interpreted to be to the left. |
 B: Cuspate fold with a tip towards the left (NE) indicating the direction of mass flow indicated also by the disaggregation of layers. |
 C: Layered pegmatites parallel to bedding. Pegmatitic melt cut across and displaced/drag folded the psammite layer. |
| Figure 7. SW of Point Ellen (about 1km?) layer parallel pegmatites are cut by metre-wide dykes trending ENE suggestging a possible two melting phases, or a melting phase straddling two deformation phases. |
 A: A younger generation of grey, granitic material forms a dyke (045/svertical) which cross-cuts early-formed migmatite characterized by banding of pegmatites and psammites.(S=350-345/50E). |
 B: Detail. Notice also how the pegmatite body in the lower right displaces blocks of psammite. |
| Figure 8. F1 intrafolial, isoclinal folds. |
 A: F1 tight fold with ~north trending axis and folded by ENE gentle F2 folds (S0=180/54W, fold axis F1=195/28) |
 B: Isoclinal F1 fold (upright, horizontal fold) with axial planar veinlets. |
| Figure 9. Schematic down plunge section of fold collapse. This is mostly based on the documentation of the structures above, but some of the interpretations are based on my experience elsewhere. |
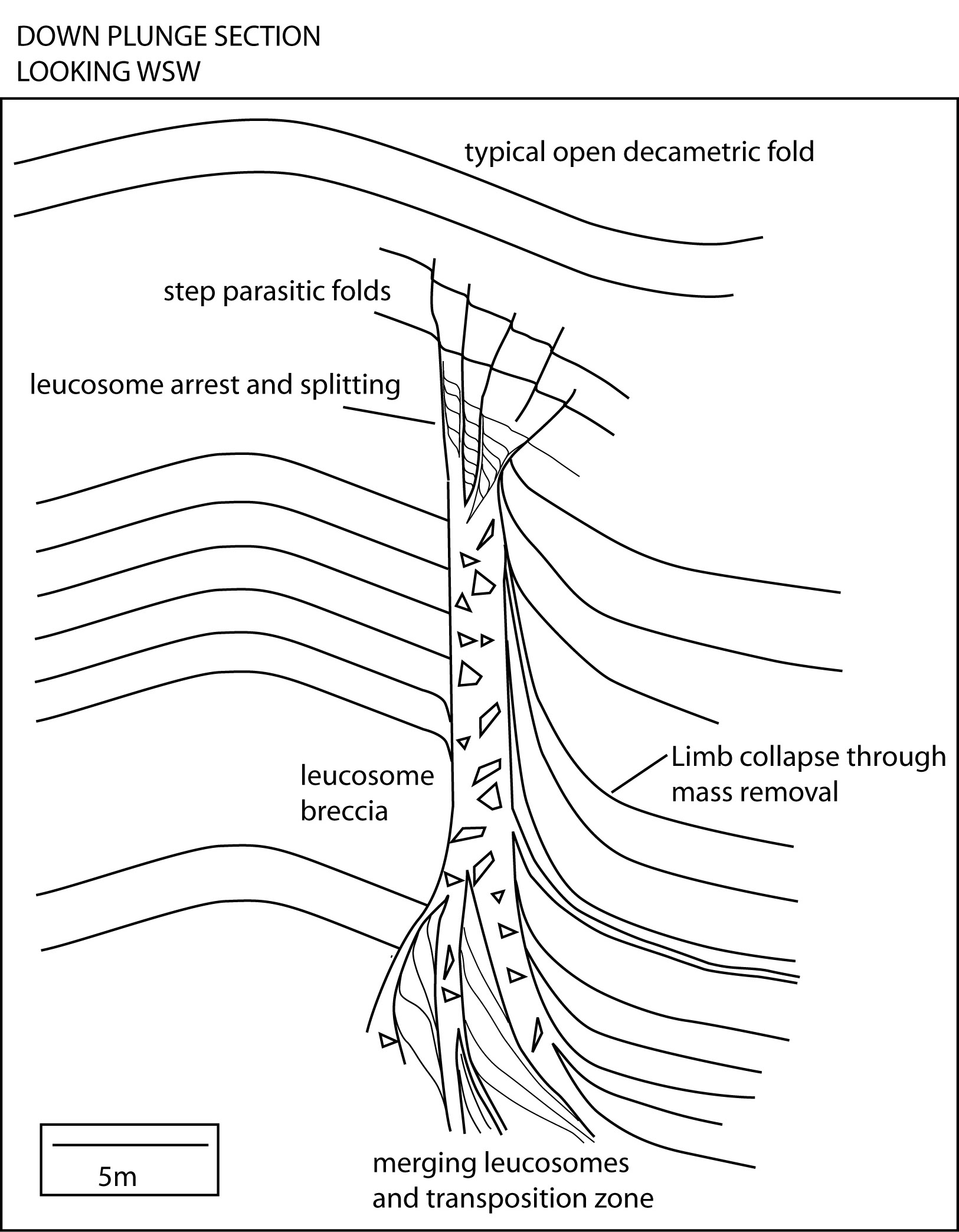
|